Acidi grassi e nutrizione dei lieviti nella fermentazione: il ruolo nascosto nella qualità del vino
In enologia si parla sempre più spesso di nutrizione del lievito, un aspetto cruciale per garantire fermentazioni regolari e vini di qualità. Gran parte della ricerca e della pratica si è finora concentrata sull'apporto di azoto, in particolare dell’azoto α-amminico, con ottimi risultati nell’ottimizzazione della vitalità e dell'attività fermentativa dei lieviti. Tuttavia, c’è un altro elemento, meno esplorato ma altrettanto strategico, che inizia ad attirare l’attenzione degli esperti: gli acidi grassi.
Sono davvero essenziali per il lievito? Oppure la loro funzione può essere compensata da meccanismi endogeni? In questo articolo esploriamo il ruolo degli acidi grassi a lunga catena nella biologia del lievito e nella dinamica fermentativa del mosto. Analizzeremo la loro sintesi, il coinvolgimento nei processi cellulari e il loro impatto concreto sulla produzione di acido acetico, un parametro chiave nella qualità finale del vino. Il tutto con uno sguardo critico alle implicazioni pratiche in cantina e alle prospettive future della ricerca.
La nutrizione del lievito: stato dell’arte in enologia
Negli ultimi tempi in enologia si è fatto molto nel campo della nutrizione del lievito, infatti l’attivante enologico organico è oggi conosciuto ed utilizzato ampiamente; grande attenzione è stata data in soprattutto all’azoto α-amminico, al fine di offrire all’enologo la possibilità di incrementare la frazione amminoacidica del mosto. Oggi molti attivanti in commercio riescono a produrre un incremento dell’azoto α-amminico nell’ordine di 20 – 30 mg/L ai dosaggi di 50 – 70 g/hL forniti all’inoculo del “pied de cuve”: questo azoto permette di migliorare la sintesi dei pool enzimatici, degli acidi nucleici e delle membrane plasmatiche del lievito.
Acidi grassi e membrana del lievito: una funzione strutturale
cruciale
Minore attenzione è stata posta agli acidi grassi a lunga catena (C16 e C18) che vanno a comporre le code alifatiche dei diacilgliceroli, elementi fondamentali della membrana fosfolipidica. Ad oggi l’unica fonte di acidi grassi C16 e C18 può essere considerata la scorza di lievito, che in effetti dà un certo aiuto nella gestione dei mosti bianchi molto chiarificati (NTU < 50): non è però esattamente quantificabile l’incremento in acidi grassi che il suo impiego può determinare.
A differenza degli amminoacidi, gli acidi grassi non sono strettamente essenziali in quanto il lievito può sintetizzarli partendo dal glucosio, via acido piruvico, sino al palmitato (o acido palmitico).
Il metabolismo lipidico del lievito
L’acido palmitico è un acido carbossilico saturo (C16:0) in cui vi sono 16 atomi di carbonio, con il carbonio C1 coinvolto nel gruppo funzionale carbossilico e gli altri 15 atomi di carbonio a comporre la coda alifatica; lo 0 dopo i due punti indica che non vi sono doppi legami nella catena, perciò ogni carbonio in essa è legato a due carboni adiacenti e a 2 idrogeni laterali, tranne il C16 che è legato a 3 idrogeni.
Dall’acido palmitico si ottiene per desaturazione via NADP+ l’acido palmitoleico (C16:1, Δ9), un acido grasso monoinsaturo con insaturazione (doppio legame) in posizione C9. L’acido palmitoleico è anche un omega 7 (ω-7): questa dicitura si trova spesso in ambito nutrizionale e conta le insaturazioni partendo dal carbonio terminale, in questo caso il C16 (ω-1).

Per elongazione dell’acido palmitico si ottiene l’acido stearico (C18:0); infine, per desaturazione via NADP+ dell’acido stearico si ottiene l’acido oleico (C18:1, Δ9), insaturo in C9, ovvero un omega 9 (ω-9); con una successiva desaturazione in C12 si ottiene l’acido linoleico, (C18:2, Δ9,12) ovvero un omega 6 (ω-6).
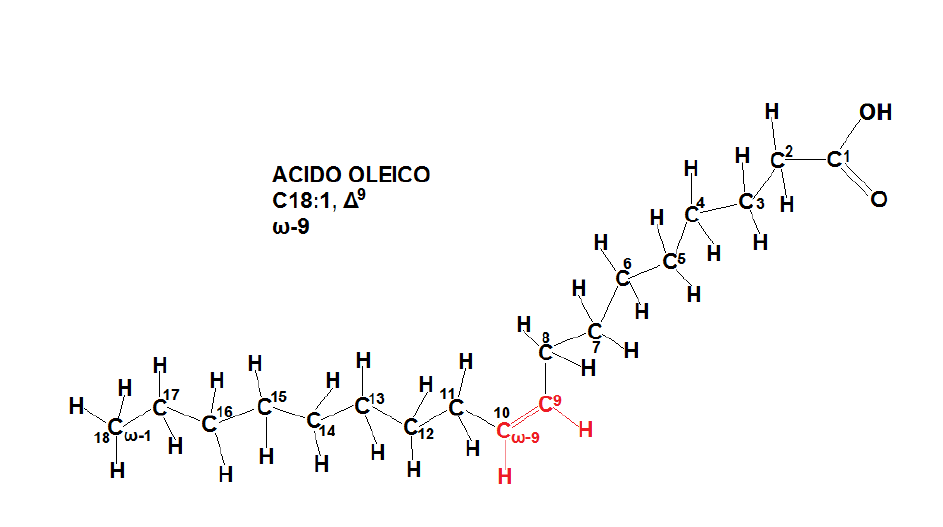

Questi 5 acidi grassi (palmitico, stearico, palmitoleico, oleico, linoleico) sono i più presenti nei triacilgliceroli siti nelle particelle lipidiche di Saccharomyces cerevisiae, dove svolgono il ruolo di esteri di riserva per la formazione dei fosfolipidi. La sintesi di una singola molecola di palmitato richiede:
l’impiego di una molecola di acetil coenzima A, che deriva dal piruvato ed è il punto di partenza per la sintesi del palmitato
l’impiego di 7 molecole di malonil coenzima A (che derivano dall’acetil coenzima A) necessarie all’elongazione dell’acido grasso da 2 a 16 atomi di carbonio
la defosforilazione di 7 molecole di ATP che servono alla sintesi del malonil coenzima A partendo da acetil coenzima A e ione bicarbonato
l’ossidazione di 14 molecole di NADPH,H+ per ridurre il numero di ossidazione degli atomi di carbonio fornendo in tutto 28 elettroni.
(fonte: Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Autori: Gianfranca Carta, Elisabetta Murru, Sebastiano Banni, Claudia Manca).
Un successivo step metabolico è la desaturazione del palmitato e dello stearato. Questo processo precede la sottrazione di due idrogeni (in posizione 9, Δ9) dalla catena alifatica dei due acidi grassi saturi. Tale deidrogenazione è svolta dal NADP+, con formazione di NADPH,H+; il NADPH,H+ deve essere poi riossidato a NADP+ per azione di una ½ O2 che sottrae i due idrogeni liberando una molecola di H20.
Nelle condizioni fermentative l’O2 (ossigeno molecolare) è quasi assente dal mezzo e costituisce quindi un fattore che limita la sintesi degli acidi grassi monoinsaturi; ne consegue la difficoltà oggettiva di ottenere la giusta quantità di acido palmitoleico e di acido oleico e il rischio di una carenza di questi acidi grassi monoinsaturi nelle catene alifatiche dei fosfolipidi. È inoltre ipotizzabile che l’acetil coenzima A, in una situazione di stallo nella sintesi degli acidi grassi monoinsaturi possa andare incontro ad una idrolisi con liberazione di acido acetico.
Impatto degli acidi grassi sulla fermentazione e sul vino
L’insieme di queste problematiche ha portato il mondo della ricerca ad interrogarsi se un mezzo arricchito in acidi grassi insaturi possa permettere al lievito una migliore performance con minore produzione di acido acetico.
Effettivamente, a livello sperimentale, l’arricchimento di mezzi fermentativi sintetici con acido oleico e linolenico ha dato una risposta positiva da parte del lievito, con una maggiore rapidità fermentativa e un minor tenore in acidità volatile: 0,24 g/l di acido acetico prodotto in presenza di 1,5 g/L di acidi grassi monoinsaturi esogeni contro 0,60 g/L di acido acetico prodotto in assenza di acidi grassi monoinsaturi esogeni. (fonte: Factors affecting acetic acid production by yeasts in strongly clarified grape musts.
Autori: E. Garcia Moruno, C. Delfini, E. Pessione, C. Giunta, Istituto Sperimentale per l’Enologia di Asti; Dipartimento Biologia Animale, Università di Torino).
Oltre l’azoto: verso una nuova analisi del mosto
Oggi è possibile eseguire analisi dettagliate sulla dotazione azotata del mosto, mentre è meno facile ottenere informazioni sulla dotazione in acidi grassi, diacilgliceroli e triacilgliceroli.
Il mio augurio è che lo studio degli acidi grassi e degli esteri del glicerolo possa avere in futuro una crescita nel settore enologico; penso inoltre possa essere utile che la ricerca enologica si soffermi sugli steroli, i coenzimi ossidoriduttivi (NAD+, NADP+) e il coenzima A, con il fine ultimo di fornire soluzioni adeguate per condurre senza difficoltà le fermentazioni di mosti poveri in questi composti.
Vuoi migliorare la gestione delle fermentazioni in cantina? Approfondisci il ruolo degli acidi grassi e applica soluzioni nutrizionali avanzate per ottenere vini più stabili, puliti e di alta qualità.
Contattaci per consulenze o strumenti innovativi dedicati alla nutrizione del lievito!